 Etsuro Takagi’s Website
Etsuro Takagi’s Website
研究概要
種子食性昆虫は樹木の種子生産に対して極めて大きい負の影響を与えます.たとえば,雌成虫(母親)による産卵場所の選択は,幼虫のパフォーマンスや生死を左右します.そのため母親は,適応度を高めるために好適な資源(e.g. 受精種子 etc.)への選択的な産卵や,重複産卵を回避すると考えられます.
そこで,モチノキとその種子を加害するモチノキタネオナガコバチを対象として,種子食性昆虫による産卵場所選択様式を明らかにすることを目的としました.

2008年と2009年に行った,野外調査,室内実験,野外実験の結果から,モチノキタネオナガコバチは,モチノキの受精種子に選択的に産卵していることが明らかになりました.
針葉樹の種子に寄生する近縁のコバチには,未受精の種子を発育させる(寄主操作)ものがいます.選択的産卵と寄主操作は,寄主植物の受精の時期と,種子食性昆虫の産卵時期の関係によって決まるという仮説が,本研究による支持されました.
また,2008年~2012年の5年にわたる野外調査からは,モチノキタネオナガコバチが,モチノキの種子内の既産卵数を識別し,重複産卵を避けていることが示唆されました.


現在,SEMによる形態観察や,産卵行動の観察によって,こうした卵分布様式になるメカニズムの解明を行っています.
Takagi, E. & Togashi, K. (2013) Distribution patterns of Macrodasyceras hirsutum eggs among Ilex integra seeds and berries. Canadian Entomologist, 145, 639-646.
Takagi, E., Iguchi, K., Suzuki, M. & Togashi, K. (2010) Selective oviposition in fertilized seed of Ilex integra by the wasp Macrodasyceras hirsutum (Hymenoptera: Torymidae). European Journal of Entomology, 107, 197-202.
生物多様性は,種の多様性だけでなく,生物間相互作用によって生み出されています.最近になって,植物の形質を介した間接効果が注目され始めました.こうした多種間相互作用は,直接的な生物間相互作用だけでなく,間接的な生物間相互作用を生み出します.この間接的な生物間相互作用は,直接的な2種間相互作用で予想される結果とは生態学的・進化学的に異なる結果を生み出すことが明らかになりつつあります.
しかし,果実や種子には多くの生物間相互作用があるにも関わらず,その形質を介した間接的な生物間相互作用に関する研究は行われてきませんでした.
そこで,東アジアを代表する鳥散布樹種と,その種子を加害するモチノキタネオナガコバチを用いて,樹木-種子食性昆虫-果実食性鳥類間相互作用系を明らかにすることを目的としました.


2008年~2011年に行った,野外調査と野外実験の結果から,モチノキタネオナガコバチは,モチノキの果実が赤色になることを阻害(果実色操作)していることが明らかになりました.
またこの果実色操作は,モチノキと鳥類間の相利共生のレベルを低下させることが示唆されました.

Takagi, E., Iguchi, K., Suzuki, M. & Togashi, K. (2012) A seed parasitoid wasp prevents berries from changing their colour, reducing their attractiveness to frugivorous birds. Ecological Entomology, 37, 99-107.
植食性昆虫の寄主シフトと特殊化は,生物多様性を創出する要因の一つです.
たとえば単食性昆虫は,これまで分布していなかった地域に侵入したときに,新しい餌を寄主にすることがあります.こうした,新しい餌を寄主に組み込む能力は,地理的な分布拡大を可能にするとともに,地域間での寄主の違いを招き,種分化を促進することがあります.
このような場合,各地域では単食性でも,他の地域と合わせると狭食性(数種を餌とする)となります.
そこで,モチノキのスペシャリスト種子捕食者であるモチノキタネオナガコバチがモチノキの近縁種に寄生するかを実験しました.
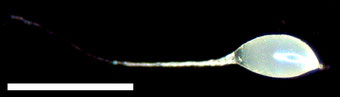
モチノキタネオナガコバチの雌成虫に,タラヨウ(モチノキの近縁種)のみを与えたところ,モチノキタネオナガコバチはタラヨウの種子に産卵しました.また,幼虫は終齢まで発育しました.
一方,野外のモチノキとタラヨウが生育している地域では,いまのところまだタラヨウへの加害は見つかっていません.
このことから,モチノキタネオナガコバチは,モチノキがない場合には,近縁種のタラヨウを利用することが明らかになりました.
また,北アフリカから東日本にかけて広く分布しているツヤクロマルカスミカメについて,北方遺存種(氷河期の分布から取り残された場所に生息している種)が多く生息する長野県菅平高原において,その寄主植物を調査しました.

ツヤクロマルカスミカメは,ユーラシア大陸ではホタルブクロ属(キキョウ科)を加害します.しかし菅平高原では,ツリガネニンジン(ツリガネニンジン属)を加害し,ホタルブクロ属は加害しませんでした.
他の地域の寄主植物に関しては現在調査中ですが,もしかしたら菅平高原で寄主植物のシフトによる種分化が起きているのかもしれません.
Takagi, E. (2014) Herbivory by Strongylocoris leucocephalus (Hemiptera: Miridae) on a novel host plant Adenophora triphylla var. japonica in Japan. Journal of Asia-Pacific Entomology, 17, 499-503.
Takagi, E. & Togashi, K. (2013) Oviposition of the seed parasitoid wasp Macrodasyceras hirsutum (Hymenoptera: Torymidae) into seeds of nonhost tree Ilex latifolia. Journal of Entomological Research Society, 15(2), 17-20.